MECANISME MOLECULAIRE DU TRANSPORT NUCLEAIREd'après le cours du Dr NEHRBASS
INTRODUCTION L’enveloppe nucléaire renferme l’ADN et défini le compartiment nucléaire. Elle est formé de 2 membranes concentriques qui se continuent avec le réticulum endoplasmique. Bien que la membrane interne et externe du noyau soit continu, les deux membrane ont une composition en protéines différentes. La membrane interne est entouré par la membrane externe qui ressemble très fortement à la membrane du R.E.R. . Comme celle-ci, elle est parsemée de ribosomes engagés dans la synthèse protéique. Les protéines synthétisées sur ces ribosomes sont transportées dans l’espace compris entre les membranes interne et externe du noyau (l’espace périnucléaire), qui est continu avec la lumière du R.E.
Le traffic bidirectionnel apparaît continu entre le cytosol et le noyau. La plupart des protéines qui ont une fonction dans le noyau - incluant les histones, les ADN et ARN polymérases, les protéines régulateurs de gènes, les protéines permettant les transformations physicochimiques (le "processing" des anglo-saxons) de l’ARN - sont sélectivement importé du cytosol où elles ont été synthétisé vers l’intérieur du compartiment nucléaire. Dans le même temps, les ARNt et ARNm qui ont été synthétisés dans le noyau sont eux exporté vers le cytosol. Comme le processus d’importation, le processus d’exportation est sélectif : les ARNm, par exemple, sont exportés seulement après qu’ils aient été modifié correctement par un « processing » de cet ARN dans le noyau. Dans certain cas, le processus de transport est complexe : les protéines ribosomales, par exemple, sont synthétisées dans le cytosol, importées dans le noyau-où elles sont assemblées en particules avec des ARNr nouvellement synthétisés- et ensuite ré-exportées vers le cytosol en tant que sous-unité ribosomale. Chacune de ces étapes implique un transport sélectif au travers de la membrane nucléaire.
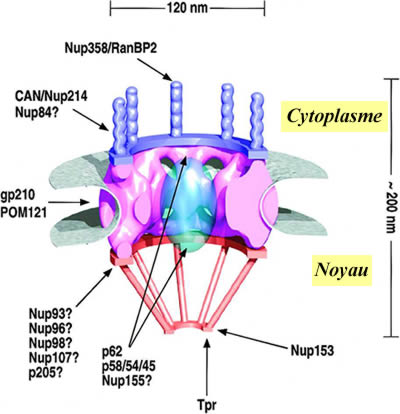
I- LES PORES NUCLEAIRES L’enveloppe nucléaire chez tous les eucaryotes est perforées par des pores nucléaires. Chaque pore est formé par une structure élaboré appelé le complexe du pore nucléaire, qui semble être composé d’au moins 100 protéines différentes, arrangées selon une symétrie octagonale.
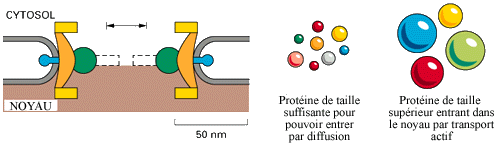
Chaque complexe contient un ou plusieurs canaux aqueux ouverts à travers lesquels des molécules solubles dans l’eau peuvent traverser la membrane nucléaire par diffusion passive, mais seulement si leurs taille sont suffisamment petites : les molécules inférieurs à 5000 Da diffuse si rapidement que l’on peut considérer que l’enveloppe nucléaire est totalement perméable à celles-ci.
Des études ont montré que la taille des canaux est d’environ 9nm de diamètre pour 15nm de longueur, occupant donc un faible volume du complexe de pore nucléaire. Mais un protéine de taille supérieur, par exemple une protéine de 44 000 Da met environ 40mn pour être équilibré entre le cytosol et le noyau. Et comment pourrais passer une protéine de 100 à 200 000 Da ? Il y a donc nécessité d’un transport actif pour faire traverser l’enveloppe nucléaire à des protéines d’une taille relativement importante.
II- LE NLS (NUCLEAR LOCALISATION SIGNAL) En général, plus le noyau est actif durant la transcription, plus son enveloppe contient un nombre important de complexe de pore nucléaire. L’enveloppe nucléaire d’une cellule de mamifère type contient entre 3000 et 4000 NPC (Nuclear Pore Complex). Si la cellule synthétise de l’ADN, elle a besoin d’importer environ 106 molécules d’histones du cytosol toute les 3 minutes, pour compacter l’ADN néosynthétisé en chromatine. Cela signifie que chaque NPC doit transporter environ 100 molécules d’histone par minute. Si la cellule est en croissance rapide, chaque NPC doit également transporter chaque minute 6 sous-unités ribosomales nouvellement assemblées du noyau, où elles sont produites, vers le cytoplasme, où elles sont utilisées. Et ceci n’est qu’une faible part du traffic total qui transite à travers les pores nucléaires. Lorsque les protéines nucléaires sont expérimentalement
extraites du noyau et micro-injecté dans le cytoplasme, une grande partie
d’entre-elles se réaccumulent dans le noyau. La sélectivité de
l’importation de ces protéines résident dans des résidus appelé
Nuclear Localisation Signal (NLS). Ce signal est présent uniquement
chez les protéines nucléaires. Il peut être localisé n’importe
où dans le séquence de la protéine, et consiste généralement en une
courte séquence (souvent entre 4 et 8 acides aminés) qui varie selon
les protéines nucléaires mais souvent riche en acide aminés chargé positivement
(Lysine, Arginine) et contenant parfois des prolines :
III- STRUCTURE DES PORES NUCLEAIRES Le NPC est une structure supra-moléculaire d’environ 125 million Da chez les vertébré et de 65 million Da chez la levure.
Les NPC s’assemble après chaque cycle cellulaire par en procédant par étape d’assemblage de sous-complexe pré-formé. Ils sont composés de 30 à 50 protéines différentes référencé comme des nucléoporines (Nups), chacune de ces nucléoporines pouvant être présente en 8 ou plus copies par NPC. Une famille de Nups contenant un nombre important de peptide FG répété est distribuée à travers tout le NPC.
Quelques exemples
de protéine et de catégorie de protéines du NPC :
IV- LE TRANSPORT NUCLEAIRE ACTIF A - Fixation (Docking) de la protéine possédant un NLS sur la face cytoplasmique du NPC Cette étape est indépendante
de l'énergie et de la température. Elle fait intervenir deux type de
protéines : les karyopherines a et b
et la protéine RanGTP. Les karyophérines a (cf. structure) s'accroche au NLS et augmente sont affinité lorsqu'une karyophérines b (cf. structure) se fixe sur une a. Les sous-unités de karyophérines coopèrent pour fixer les séquences répétées FG des nucléoporines : les karyophérines b fixent les nucléoporines sur les répitions peptidique FG. Treize des trentes nucléoporines possédant une séquence peptidique répété FG, les karyophérines peuvent charger les protéines substrats sur de multiple site le long du NPC ( Il existe probablement aux alentours de 200 sites de fixation aux karyophérines). B - Import dans le noyau et désassemblage du complexe d'importation Une fois fixé, le complexe pénètre dans le pore nucléaire. Le mécanisme de translocation semble inconnue. Arrivé dans le noyau, RanGTP se fixe au karyophérines b et cause la dissociation du complexe et de la nucléoporine par un mécanisme allostérique.
L'exportation se fait selon un procédé similaire, à celà près que l'on n'a plus affaire à des importines mais à des exportines et que la molécule responsable de la dissociation du complexe est RanGDP
Le transport nucléaire est donc une suite d'importation et d'exportation de protéine gouvernée par un gradient de RanGTP, faiblement concentré dans le cytoplasme et fortement concentré dans le noyau. Et pour maintenir ce niveau de RanGTP dans le noyau, une enzyme, RCCI ou RanGEF (cf. structure), convertie le RanGDP en RanGTP, tandis que dans le cytoplasme, une autre enzyme, RanGAP (cf. structure), convertie le RanGTP en RanGDP.
Schéma Récapitulatif BIBLIOGRAPHIE: Revues : Articles : J.E. Hinshaw and R. Milligan, Cell 69:1133-1141, 1992 |